Mutation spectra (SBS-96 / SBS-192)
RIP is one of several processes that deaminate cytosine in fungal repeats. To tell
them apart you need more than a count of C→T changes — you need the sequence
context each change happened in. derip2-spectra builds the standard
single-base-substitution spectra used in mutational-signature analysis:
- SBS-96 — every substitution folded onto the pyrimidine strand, classified by its 5′ and 3′ neighbours: 6 substitution types × 16 contexts = 96 channels.
- SBS-192 — the strand-resolved form (12 types × 16 contexts) that keeps the reference base as observed on the coding strand, so strand asymmetries stay visible.
RIP (CpA → TpA) shows up as a sharp C>T peak concentrated in NCA contexts:
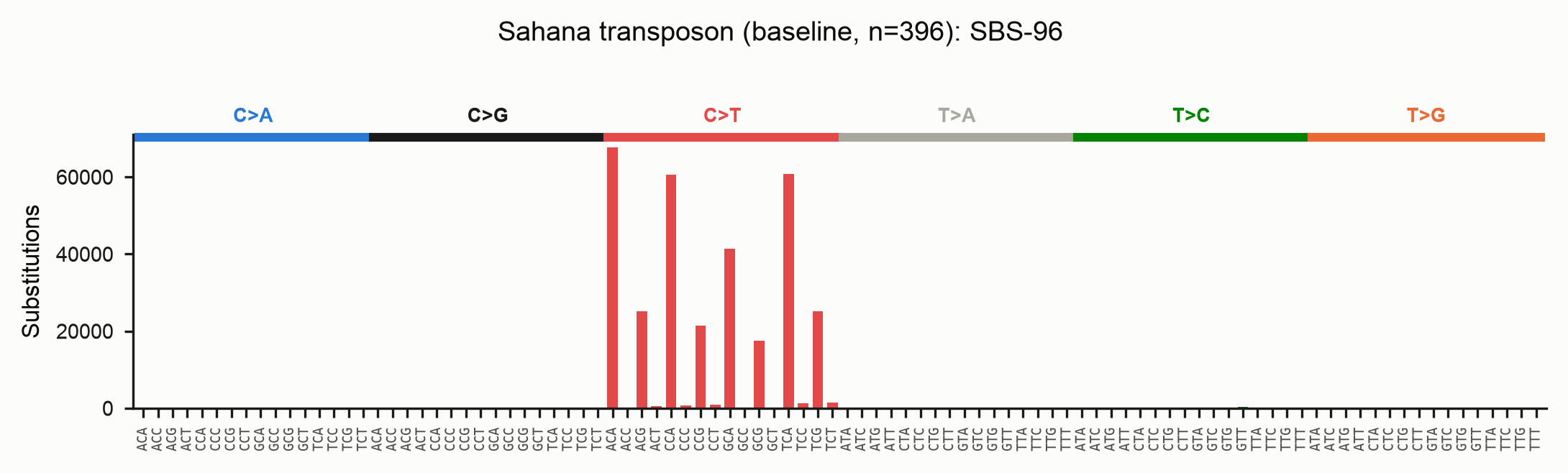
The matrices are written in SigProfiler-compliant format, so they drop straight
into SigProfilerPlotting / SigProfilerAssignment if you want to decompose them
against COSMIC signatures later.
Quick start
This writes, into out/:
| File | Contents |
|---|---|
family.SBS96.txt, family.SBS192.txt |
SigProfiler-compliant count matrices |
family_SBS96.png, family_SBS192.png |
spectrum bar plots |
family_strand_asymmetry.png |
coding- vs template-strand counts per class |
family_homoplasy.png, family_homoplasy.tsv |
recurrently-hit sites |
family_events.tsv |
one row per called substitution |
Two methods: baseline and phylogenetic
Direction and recurrence are not free from an alignment — you need an ancestor. The tool offers two ways to get one.
Baseline (--method baseline, the default)
Every sequence is compared to a single reference — deRIP2's reconstructed ancestral consensus — and each difference is one event, with its context read from that ancestor. It needs no tree and no external tools.
Its blind spot is recurrence. If the same C→T deamination struck independently on many lineages, comparing every tip to one reference records the derived state on each tip and cannot tell "one ancestral event inherited by twenty tips" from "twenty independent events". The baseline therefore over-counts homoplasic sites and reports recurrence only as a multi-hit-column proxy.
Worked example: the baseline ancestor
By default you do not have to build the ancestor yourself — derip2-spectra
reconstructs it internally by running deRIP2's correction, then compares every
sequence to it:
The ancestor it uses is deRIP2's gapped deRIP consensus: for every column,
deRIP2 finds the un-RIP'd ancestral base (a C where RIP produced T, a G
where it produced A) by looking across all copies for one that escaped RIP, and
falls back to the majority base elsewhere. This is exactly the sequence the plain
derip2 command writes to family.fasta, so the two are consistent.
If you want to see or reuse that ancestor, run derip2 first and pass it back
with --ancestor:
# 1. Reconstruct the deRIP'd ancestral consensus (writes out/family.fasta)
derip2 -i family.fasta -d out -p family
# 2. Call the spectrum against that explicit ancestor
derip2-spectra -i family.fasta --method baseline \
--ancestor out/family.fasta -d out -p family_spectrum
--ancestor accepts any FASTA whose single sequence is the same length as the
alignment (one base per column, gaps allowed). Use it when you have an
independent progenitor — a known germline element, a manually curated ancestor,
or a consensus built under different deRIP settings (e.g. derip2 --reaminate to
treat all cytosine deamination as ancestral, not just RIP-context events). The
deRIP parameters on derip2-spectra itself (--max-gaps, --reaminate,
--min-rip-like, …) tune the internal ancestor when you do not supply one.
# The same thing from Python
from derip2.derip import DeRIP
d = DeRIP('family.fasta')
d.calculate_rip()
ancestor = str(d.gapped_consensus.seq) # the deRIP'd ancestor, one base per column
result = d.calculate_spectra() # baseline spectra against that ancestor
result.sbs96 # (96, n_samples) count matrix
Reusing an ancestor already in your alignment
The plain derip2 command appends its deRIP'd consensus to the output alignment
by default, as a row whose id is its --prefix (default deRIPseq). If you feed
that alignment straight to derip2-spectra, it detects that row, uses it as the
ancestor, and excludes it from the counted sequences — no recomputation, and no
self-comparison inflating the counts:
# 1. deRIP the family with the default prefix; this writes
# out/deRIPseq_alignment.fasta with a "deRIPseq" consensus row appended
derip2 -i family.fasta -d out
# 2. Spectra reuse that row automatically (logged: "Using pre-computed reference
# 'deRIPseq' from MSA; excluding it from counted sequences")
derip2-spectra -i out/deRIPseq_alignment.fasta -d out -p family_spectrum
Because the appended row takes derip2's --prefix, the spectra default
--reference-tag deRIPseq matches a default derip2 run. If you deRIP'd with a
different prefix (or curated the row by hand), name the tag to match — it is an
exact id match:
derip2 -i family.fasta -d out -p family # row is named "family"
derip2-spectra -i out/family_alignment.fasta --reference-tag family -d out -p fam_spec
Precedence is explicit: an --ancestor FILE always wins over an in-alignment row,
which in turn wins over recomputing the consensus.
Input must be unambiguous DNA
Alignments may contain only A/C/G/T/- (upper or lower case — soft-masking is
normalised before analysis). Degenerate IUPAC characters (N, R, Y, …)
are rejected with an error naming the offending character and its location,
rather than being silently coerced to gaps as they were previously.
Phylogenetic (--method phylo)
The rigorous path reconstructs ancestral sequences at every internal node of a tree (IQ-TREE marginal ASR) and walks every parent→child branch, logging each substitution as an independent event with its context read from the parent sequence — the state at the moment the mutation occurred.
The difference is large and biological. On the full Sahana family (396 copies):
| Events (SBS-96) | Interpretation | |
|---|---|---|
| Baseline | 326,778 | every tip vs one reference |
| Phylogenetic | ~47,000 | independent branch events |
The baseline's extra ~280,000 "events" are shared/inherited RIP mutations counted once per descendant. The phylogenetic path assigns each to the single branch it arose on — and flags the sites that really were hit again and again:
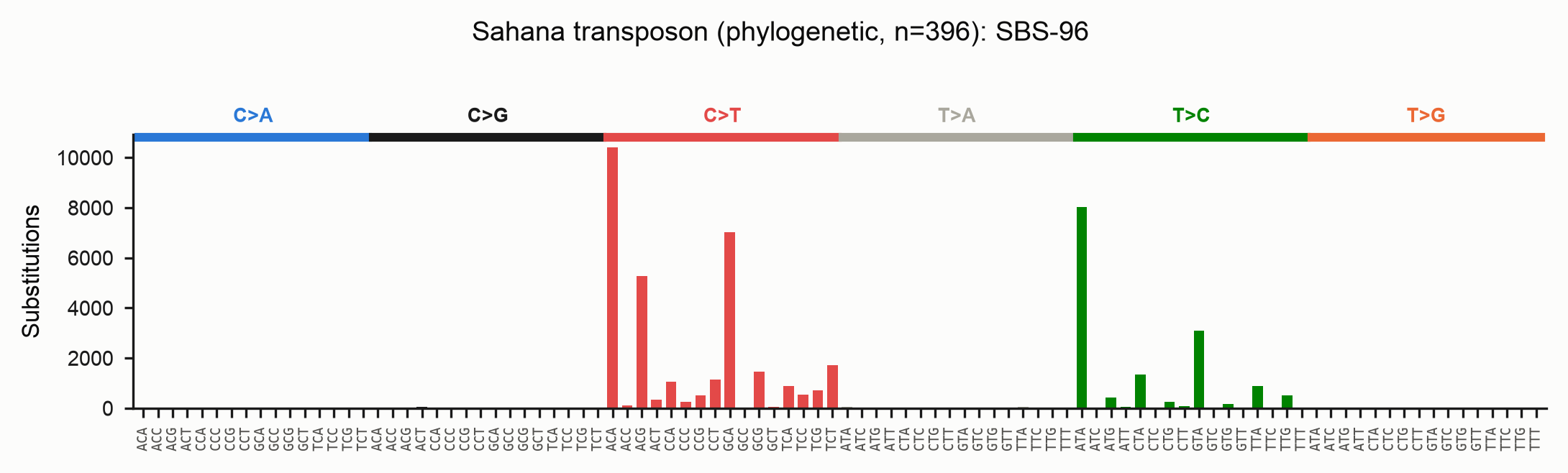
Read direction with care on heavily-RIP'd families
Notice the T>C peak in the phylogenetic spectrum, absent from the baseline.
It is largely an artefact of maximum-likelihood ancestral reconstruction,
which is biased toward the majority state. When RIP has converted most copies
of a column from C to T, IQ-TREE reconstructs the internal nodes as the
majority T, so the minority copies that retained the ancestral C read as
T>C reversals. The recurrence counting is still correct (46,952 vs
326,778 events) — this bias affects the inferred direction, not the event
count.
deRIP2's consensus-based baseline is more robust to RIP direction: its ancestor is the deRIP'd (un-RIP'd) sequence, recovered by finding the copies that escaped RIP even when they are the minority. Use the two together — the baseline for polarity, the phylogenetic path for correct recurrence — and lean on the masked-topology workflow below to keep the tree itself honest.
IQ-TREE is required for --method phylo
Install it separately (conda install -c bioconda iqtree) and make sure
iqtree3, iqtree2 or iqtree is on your PATH. ete4 (a Python
dependency) handles the tree.
# Infer the tree and reconstruct ancestors in one step
derip2-spectra -i family.fasta --method phylo -d out -p family
Supplying your own phylogeny
You will usually get a better tree from a dedicated run than from the built-in
one-shot inference. Pass any Newick tree with --tree; IQ-TREE then keeps that
topology fixed (-te) and re-estimates the model, branch lengths and ancestral
states from your alignment.
# Build a well-supported tree however you like...
iqtree3 -s family.fasta -m MFP -B 1000 -T AUTO --prefix family_tree
# ...then reconstruct ancestral states on it and call the spectrum
derip2-spectra -i family.fasta --method phylo --tree family_tree.treefile \
-d out -p family
Tip names
Tree leaf names must correspond to the FASTA sequence ids. IQ-TREE rewrites
characters outside [A-Za-z0-9._-] to _ in its output (so scf:1-9(+)
becomes scf_1-9___); deRIP2 applies the same rule to match leaves back to
sequences, so trees produced by IQ-TREE Just Work. If you supply a tree from
another tool, keep tip names to those safe characters.
Recommended: infer topology from a RIP-masked alignment
RIP mutations are convergent — the same CpA→TpA change happens independently in many copies. To a tree-builder that convergence looks like shared ancestry, so a tree built directly from RIP-riddled repeats can be pulled out of shape, grouping copies by how much they were RIP'd rather than by their true history. That distorted topology would then mis-assign the very substitutions you are trying to count.
The fix is to infer the topology from a RIP-masked alignment, then reconstruct ancestral states for the unmasked sequences on that same topology:
# 1. Mask RIP-corrected positions (degenerate IUPAC codes); no consensus appended
derip2 -i family.fasta --mask --no-append -d out -p family
# -> out/family_masked_alignment.fasta
# 2. Infer the topology from the masked alignment (RIP signal removed).
# -st DNA forces the DNA model: a heavily masked alignment carries many IUPAC
# ambiguity codes, and IQ-TREE's sequence-type auto-detection can otherwise
# fail with "Unknown sequence type".
iqtree3 -s out/family_masked_alignment.fasta -m MFP -B 1000 -T AUTO -st DNA \
--prefix out/family_masked
# 3. Reconstruct ancestral states for the UNMASKED sequences on that fixed
# topology, and call the spectrum
derip2-spectra -i family.fasta --method phylo --tree out/family_masked.treefile \
-d out -p family_spectrum
Why this works: masking removes the homoplasic RIP columns that mislead tree
search, giving a topology that reflects true descent. Step 3 then fixes that
topology (--tree) but re-derives branch lengths, the substitution model and the
ancestral sequences from the full, unmasked alignment — so the spectrum is
computed from the real substitutions while the tree shape is not an artefact of
RIP. The masked and unmasked alignments share identical tips and columns (masking
only rewrites bases in place), so the topology transfers exactly.
Same topology, unmasked ancestors
The point of --tree here is precisely that the topology comes from the
masked tree while the ancestral sequences are recomputed for the unmasked
data. Do not run IQ-TREE ancestral reconstruction on the masked alignment —
its ancestors would be masked too, and the spectrum would be blank exactly
where RIP acted.
The resulting spectrum for the full Sahana family (topology from the masked
alignment, ancestral states from the unmasked sequences) still resolves the RIP
C>T/CpA signal cleanly, now on a topology that RIP homoplasy could not distort:
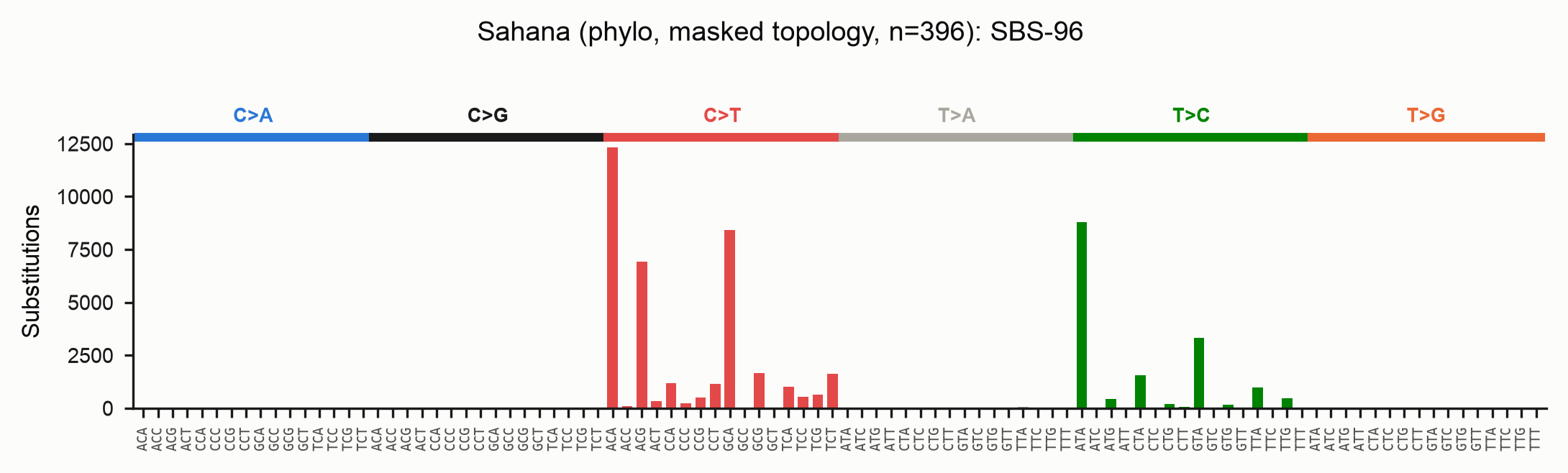
Reading the outputs
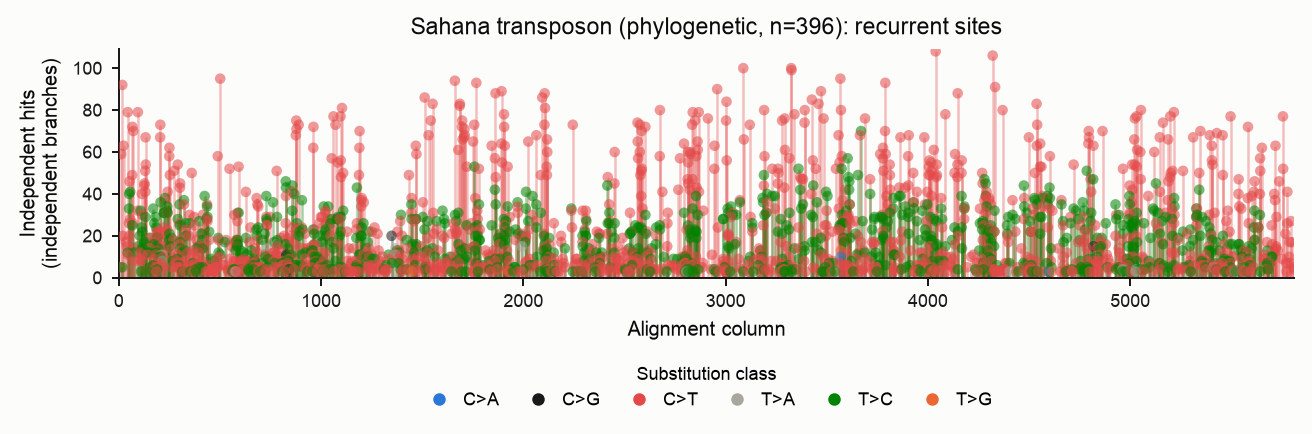
Homoplasy (recurrence)
The homoplasy report lists sites hit by the same substitution on two or more independent lineages — the explicit, measured record of recurrent deamination. Each stem is one (column, derived base); its colour is the pyrimidine-folded substitution class (see the legend) and its height is the number of independent hits. Markers are drawn semi-transparent so that stems stacked at the same column and height remain visible.
The recurrence unit differs by method, and the two plots make the point of the whole feature. Under the baseline every hit is one sequence carrying the derived state, so shared/inherited RIP makes almost every column look "recurrent" — the multi-hit-column proxy saturates:
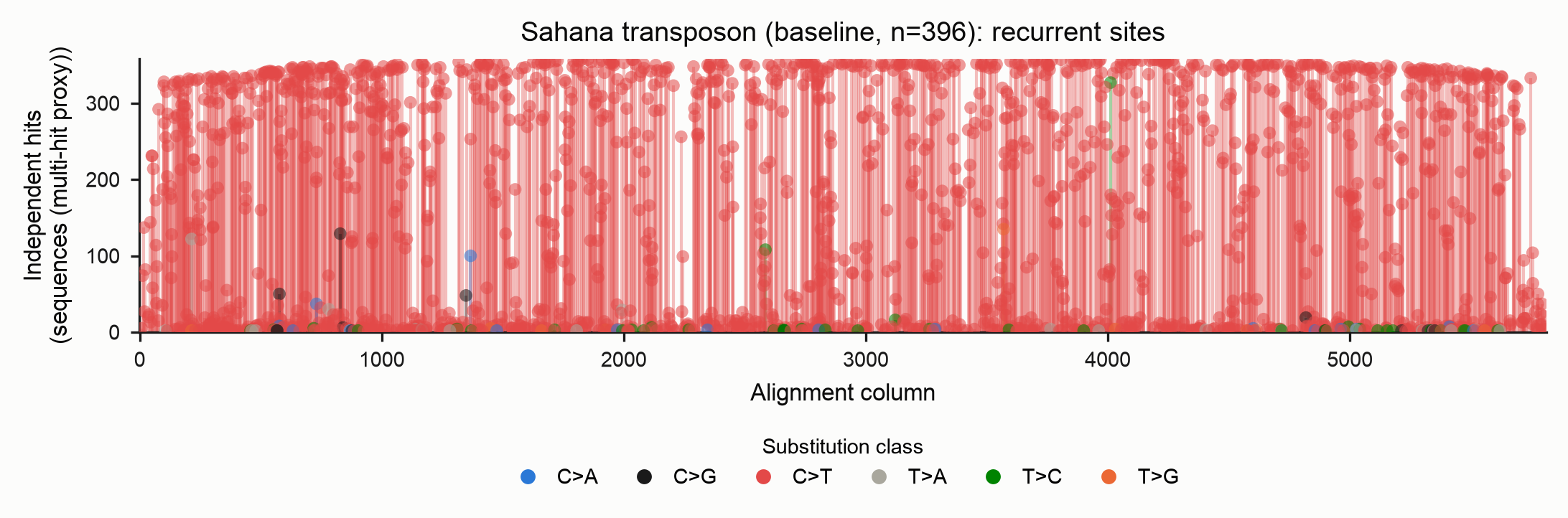
Under --method phylo each hit is an independent branch event, so only sites
that truly mutated more than once on the tree remain. The plot is far sparser and
each stem is a real recurrence (here, sites hit on ≥3 independent branches):
Strand asymmetry
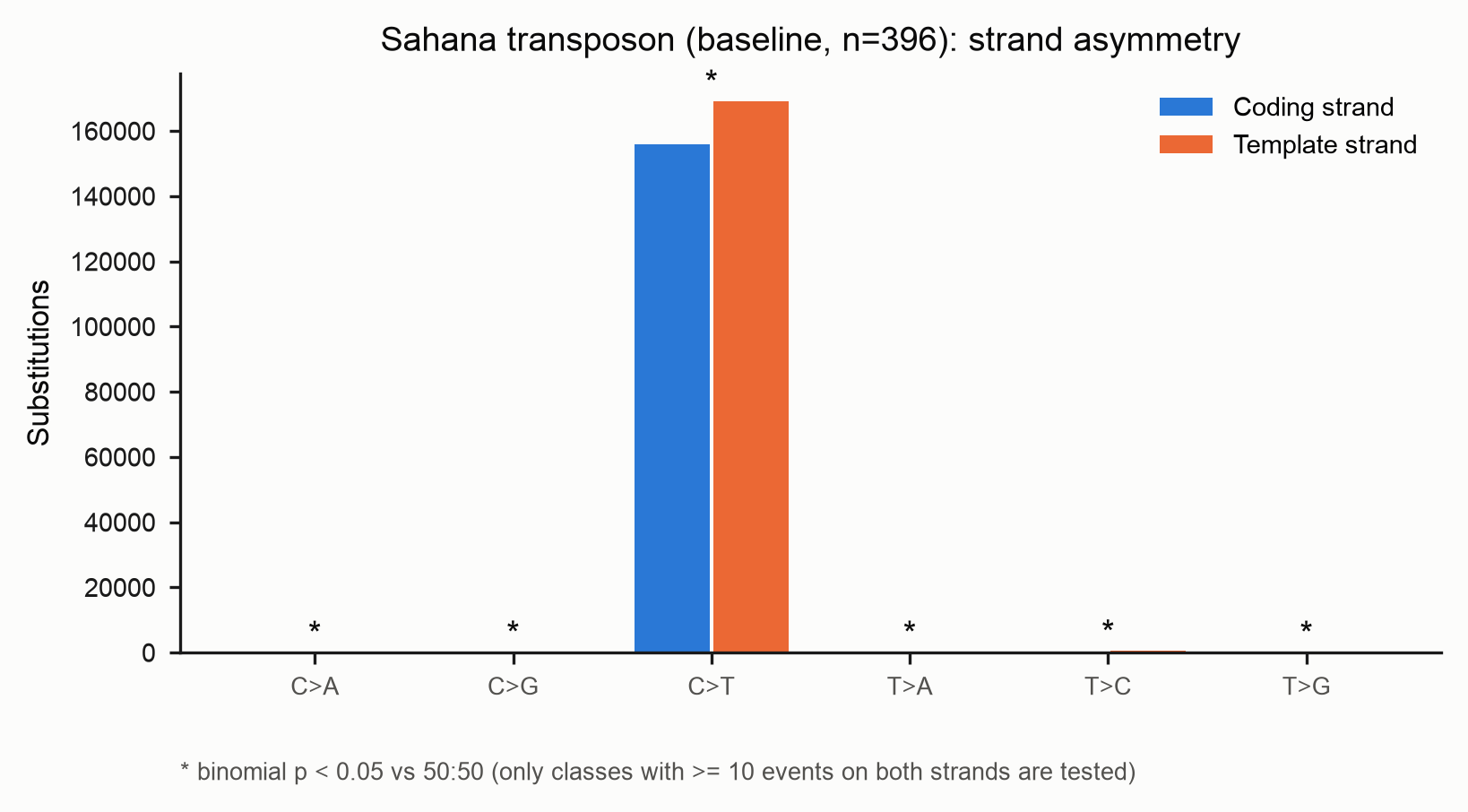
From the SBS-192 matrix, each pyrimidine class is compared to its
reverse-complement partner. For every class two bars are drawn: coding-strand
counts (blue) and template-strand counts (orange) — the legend colours the two
strands, and the substitution class is read off the x-axis. An asterisk marks a
class whose coding-vs-template split departs from an even 50:50 at a binomial
p < 0.05 (the figure footnote states this). This is where enzyme-, replication-
or transcription-linked strand biases show up.
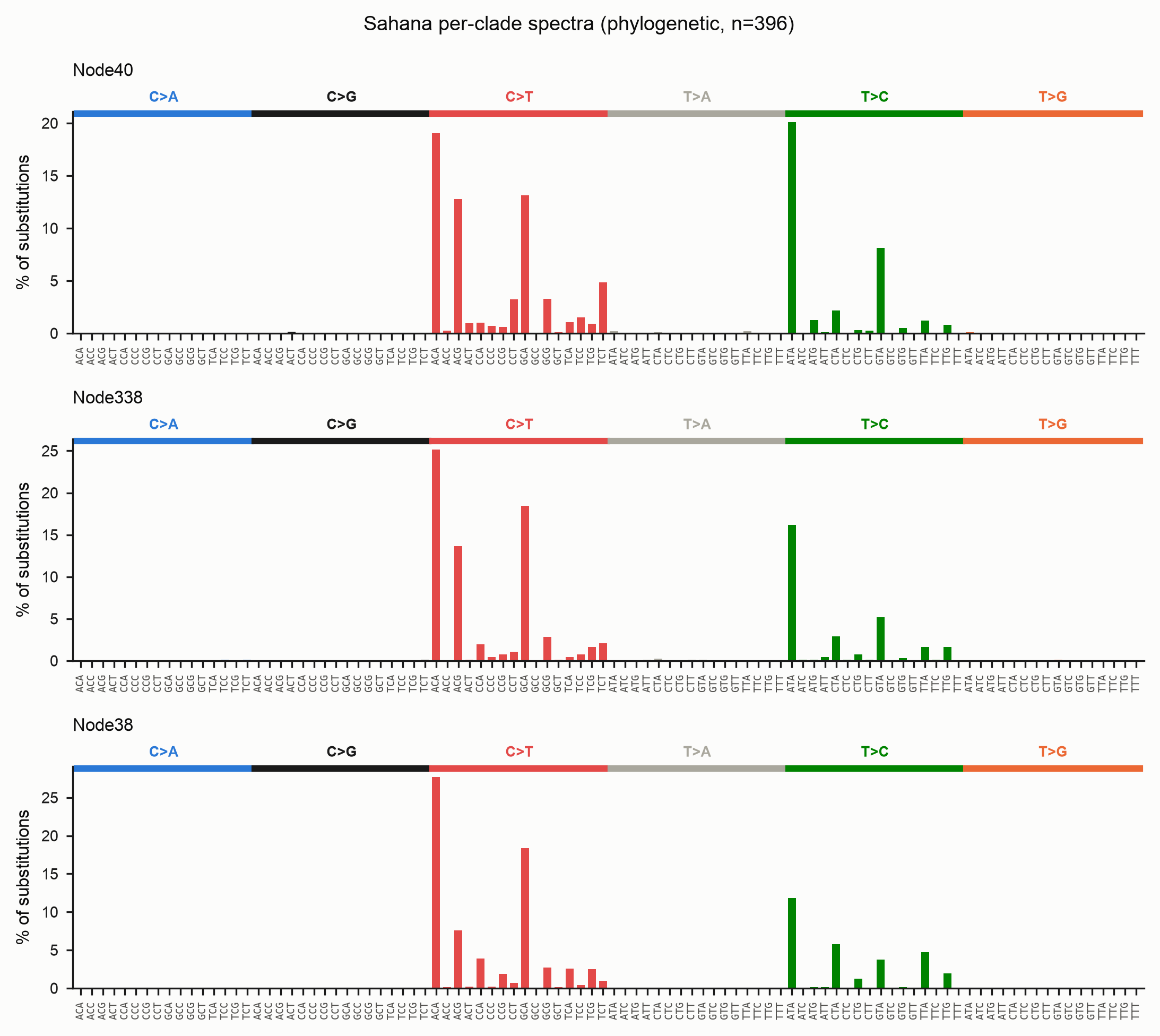
Per-lineage spectra
--partition-by clade (phylo) splits the matrices into one sample column per
subtree hanging off the root, so both the matrix files and the plots carry one
panel per lineage — useful for spotting lineage-specific deamination. Each branch
is attributed to the clade its child subtree belongs to; branches above the split
form the ancestral trunk.
--partition-by row does the analogous per-sequence split for the baseline.
Per-group spectra (species or user-defined sets)
When you already know which sequences belong together — species, populations,
sub-families — pass a two-column mapping with --groups to get one spectrum per
group. This works for both methods.
The mapping file is whitespace- or tab-separated: sequence name, then group label.
A header row and # comments are optional:
# sequence group
UNSE01000019.1:422682-431483(-) speciesA
UNSE01000019.1:709761-718562(-) speciesA
UNSE01000006.1:12043-20871(+) speciesB
Sahana_prime reference
# Baseline: one spectrum per group, each sequence compared to the deRIP ancestor
derip2-spectra -i family.fasta --groups groups.tsv -d out -p family
# Phylogenetic: a branch is attributed to a group only when its whole descendant
# clade belongs to that group (spanning branches become 'mixed')
derip2-spectra -i family.fasta --method phylo --groups groups.tsv -d out -p family
Names are matched leniently: the label file may use the original FASTA ids even
though IQ-TREE rewrites special characters in tree tip names, so the same file
works for both methods. Sequences absent from the map fall into an ungrouped
sample.
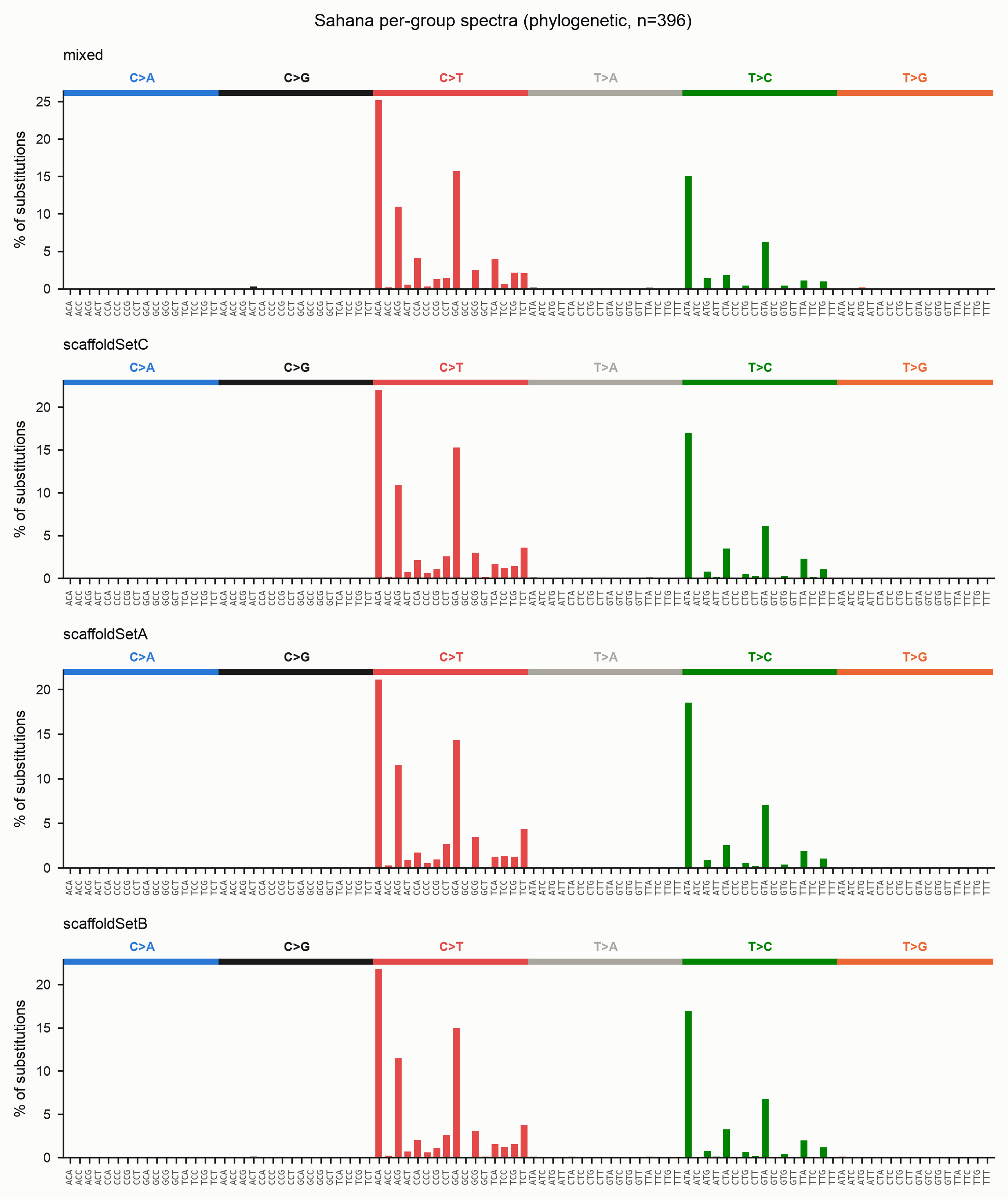
Grouping in the example
The Sahana copies have no species labels, so this figure bins them by genomic scaffold purely to illustrate the mechanic. In practice the labels would be your species or population names, and the panels would let you compare, say, RIP intensity between a methylation-competent and a methylation-deficient lineage.
Run manifest
The phylo path writes *_run_manifest.json recording the IQ-TREE version, the
model, the rooting method and the root node, node/edge counts, and — with
--root-sensitivity — the fraction of edges whose direction flips under midpoint
rooting. Directionality depends on the root, so record it.
Comparing spectra statistically
Once you have spectra for two or more groups — or two matrices from separate runs
— you will want to ask whether they actually differ. deRIP2 provides two
complementary measures in derip2.stats (no SciPy required):
- Cosine similarity — a scale-free effect size in
[0, 1]. It compares the shape of two profiles and ignores their totals, so it answers "do these look alike?". 1.0 is identical. - Chi-squared test of homogeneity — a significance test of whether the channel counts could have come from one shared distribution ("is the difference more than sampling noise?"). Its per-channel standardised residuals show which channels drive any difference, and Cramér's V is its effect size.
Read the p-value with the effect size
Spectra often carry hundreds of thousands of events, and at that sample size a chi-squared test flags even biologically trivial differences as "significant". Always read the p-value next to the cosine similarity and Cramér's V. A tiny p with cosine ≈ 1 and Cramér's V ≈ 0 means "different, but only in a way that does not matter"; a small p with low cosine and larger Cramér's V is a real shift in the spectrum.
Compare groups by label
Run with --groups (or --partition-by), then compare the sample columns of the
matrix it writes:
from derip2.spectra import read_sbs_matrix
from derip2.stats import compare_spectra, pairwise_compare
channels, samples, matrix = read_sbs_matrix('out/family.SBS96.txt')
# One pair, with the channels that differ most
a = matrix[:, samples.index('setA')]
b = matrix[:, samples.index('setB')]
res = compare_spectra(a, b, channels)
print(res['cosine_similarity'], res['pvalue'], res['cramers_v'])
for c in res['top_channels'][:5]:
print(c['channel'], c['a'], c['b'], round(c['residual'], 2))
# Every pair at once (Bonferroni-corrected), most-different first
for row in pairwise_compare(matrix, samples):
print(row['a'], row['b'], round(row['cosine_similarity'], 4),
f"p_adj={row['pvalue_adjusted']:.3g}")
For two random scaffold-based halves of the Sahana family the result is:
Cosine ≈ 1, a non-significant p and Cramér's V ≈ 0 — exactly what you expect when
both groups are shaped by the same RIP process. A methylation-competent versus
methylation-deficient pair, by contrast, would show a lower cosine, a small p and
C[C>T]G/CHG-context channels topping the residual list.
Compare two precalculated matrices
Comparing SigProfiler-format matrices from separate runs (or external tools) is the same call — read each file and pass one column from each. Both must be the same context (both SBS-96 or both SBS-192):
from derip2.spectra import read_sbs_matrix
from derip2.stats import compare_spectra
ch_a, _, mat_a = read_sbs_matrix('speciesA.SBS96.txt')
ch_b, _, mat_b = read_sbs_matrix('speciesB.SBS96.txt')
assert ch_a == ch_b # deRIP2 writes channels in canonical order, so they align
res = compare_spectra(mat_a[:, 0], mat_b[:, 0], ch_a)
print(res['cosine_similarity'], res['pvalue'], res['cramers_v'])
For a robustness check
The chi-squared test assumes each event is independent. Recurrent RIP breaks
that assumption mildly, so treat borderline p-values as a screen. When a call
is close, a permutation test — reshuffle the group labels many times and see
how often a random split reaches your observed cosine distance — is a
distribution-free alternative you can build on top of --groups.
Useful options
| Option | Purpose |
|---|---|
--sbs {96,192,both} |
which matrices/plots to produce |
--rooting {midpoint,outgroup,none} |
how to root (sets substitution direction) |
--outgroup NAME[,NAME…] |
outgroup tip(s) for --rooting outgroup |
--iqtree-model |
IQ-TREE model (default MFP) |
--threads |
IQ-TREE -T (default AUTO; pass an integer to skip its benchmark on small alignments) |
--min-prob |
drop phylo events below this parent × child ancestral posterior |
--partition-by {none,row,clade} |
pool, per-sequence, or per-clade samples |
--groups FILE |
report one spectrum per user-defined group (both methods) |
--root-sensitivity |
report how much polarity depends on the rooting choice |
--ancestor FASTA |
baseline only: call against a supplied ancestor (validated to match the alignment width) instead of the deRIP consensus |
--reference-tag ID |
baseline only: id of an ancestor row already in the alignment to reuse and exclude from counting (default deRIPseq) |
Decomposing against COSMIC signatures
Because the matrices are SigProfiler-compliant, you can fit them to reference
signatures. First render the standard SBS-96 figure (SigProfilerPlotting), then
fit COSMIC signatures with SigProfilerAssignment (both installed separately):
from SigProfilerAssignment import Analyzer as Analyze
Analyze.cosmic_fit(
samples='out/family.SBS96.txt',
output='out/cosmic',
input_type='matrix',
context_type='96',
cosmic_version=3.4,
make_plots=True,
)
Fitting the full Sahana spectrum (326,778 events) to COSMIC v3.4 gives:
| COSMIC signature | Assigned mutations | Share | Human-cancer aetiology |
|---|---|---|---|
| SBS44 | 135,625 | 41.5% | defective mismatch repair |
| SBS96 | 92,238 | 28.2% | unknown |
| SBS2 | 51,312 | 15.7% | APOBEC cytosine deamination |
| SBS1 | 47,603 | 14.6% | spontaneous 5-methyl-cytosine deamination |
The reconstruction cosine similarity is only 0.80 (a good fit is usually > 0.9). That poor fit is itself the result: RIP has no COSMIC signature, so the fitter approximates it with a blend of the human deamination signatures (SBS1, the CpG 5mC-deamination clock, and SBS2, APOBEC) plus catch-all signatures.
COSMIC signatures do not apply directly to fungi
The COSMIC reference set was derived from human cancers, on whole-genome trinucleotide opportunities. RIP and fungal methylation-driven deamination are not represented, so a decomposition like the one above is at best a loose analogy — treat the assigned signatures as "the nearest human look-alikes", not as mechanisms operating in your fungus. The low cosine similarity is the honest signal of that mismatch. The same machinery would, however, work well against a fungal-specific reference library (a custom signature matrix in the same SBS-96 format), which is the appropriate way to interpret these spectra — and building one is a natural next step for the community.
Interpret single-gene or single-family fits cautiously in any case: reference signatures assume genome-wide trinucleotide opportunities, so a normalisation caveat applies on top of the species-mismatch one.